
緒論
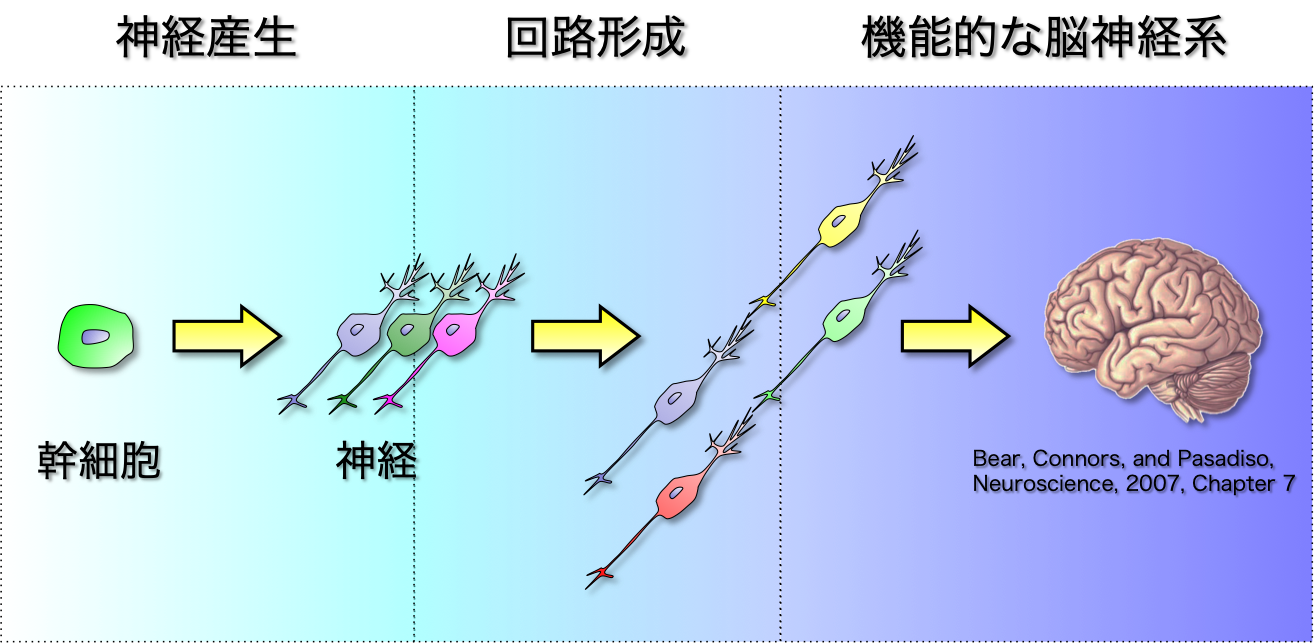
私たちの脳神経系は莫大な数の神経細胞から構成されており、それぞれが自身のパートナーと連絡することによって非常に複雑な神経回路を構築しています。 この回路によって、目、鼻、耳などの感覚器からの情報が適切に処理・伝達され、見たもの、嗅いだ匂い、聞いた音を認識しています。 このように、多数の神経細胞によるネットワークが正確に機能することによって脳機能を実現しています。それぞれの神経細胞は、初めから存在するわけではなく、 すべて神経幹細胞という1種類の細胞から生み出され、発生・成長に伴って神経回路を構築していることが知られています。
私たちの脳神経系は、いつ、どうやって、それぞれの機能を獲得するのでしょうか?
この問いかけに答えるためには、それぞれの機能を担う神経細胞が、いつ、どこで、どのようにして生まれ、いかにして正しい神経回路を形成するのか、
という脳神経系の発生メカニズムの全体像を理解することが重要です。
私たちの研究室では、研究対象としてショウジョウバエの視覚中枢に注目し脳神経系の発生メカニズムの研究を進めています。
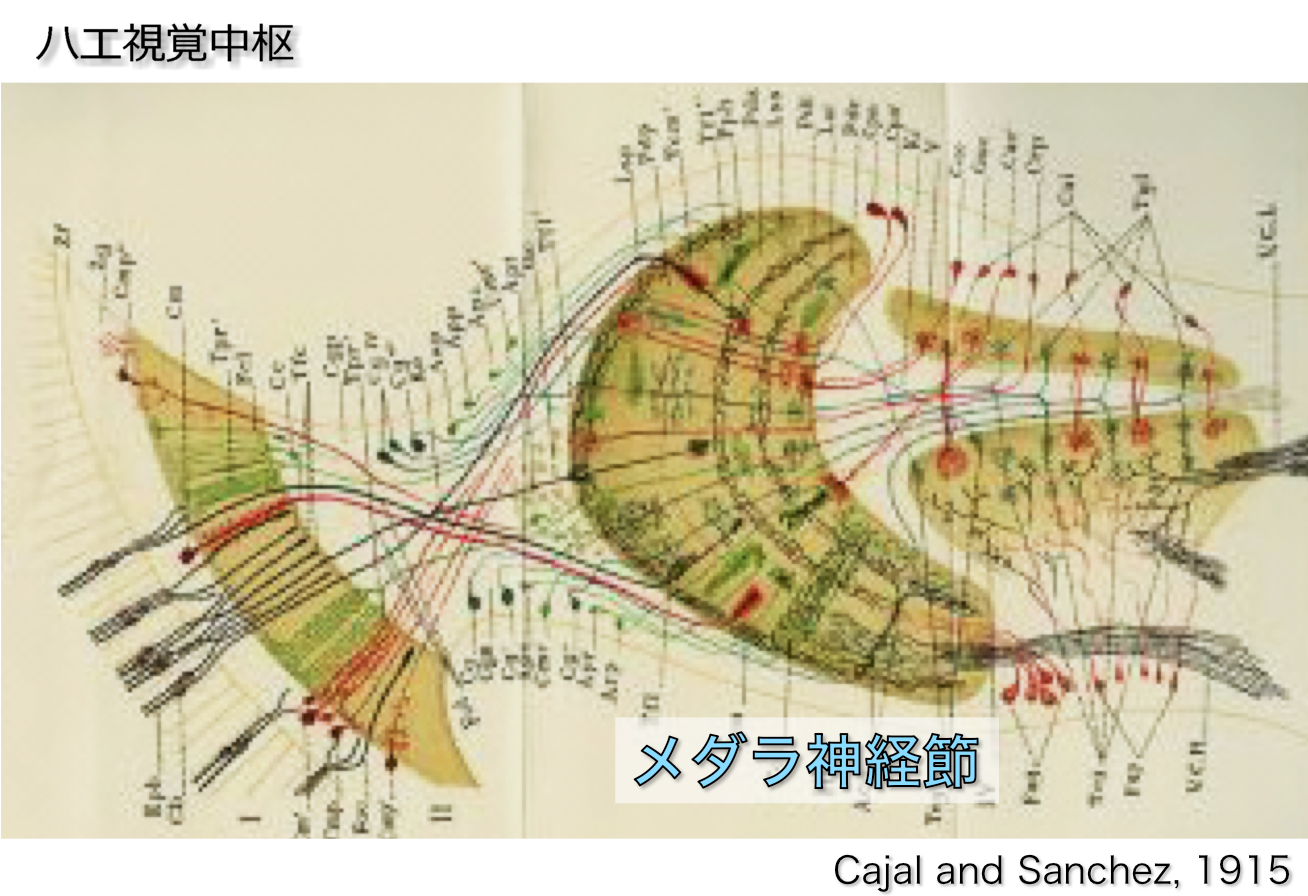
ハエの視覚中枢は網膜、ラミナ神経節、メダラ神経節、ロビュラ神経節の4つの領域から構成されています。
その中で最大のメダラ神経節は、層構造・カラム構造など哺乳類の大脳と非常によく似た構造をしており、神経細胞の総数・種類ともに哺乳類大脳と比較してはるかに少なく解析モデルとして非常に適しています。
ハエはライフサイクルが短く、さらに遺伝学的な解析手法が発展しており、他の生物では困難な解析でも比較的容易に実施できるという大きな利点を持っています。
神経系の細胞多様性を生み出す仕組み
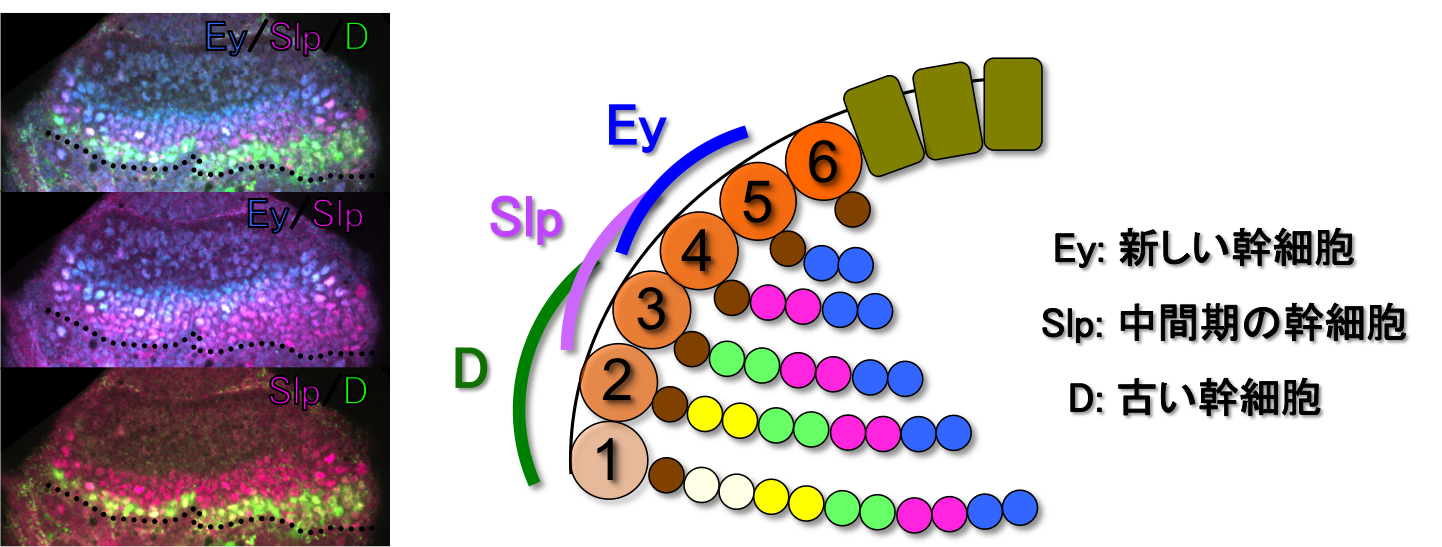
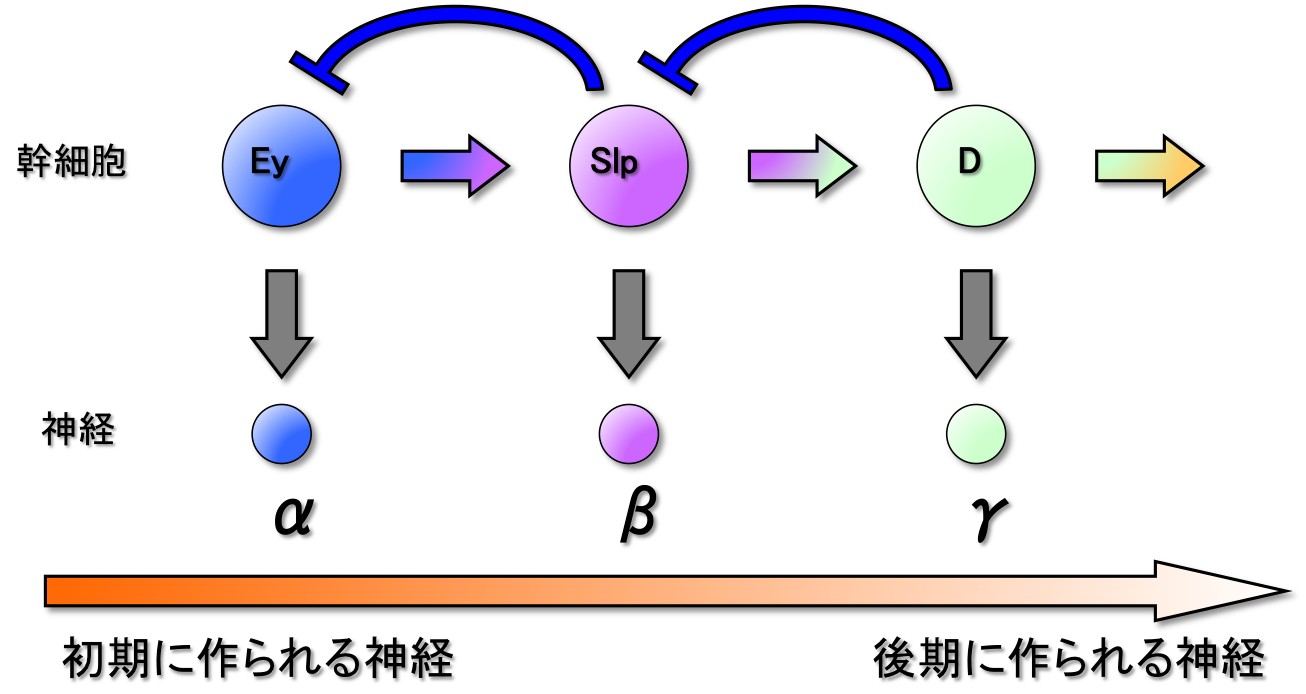
私たちの脳神経系は、小型から大型、単純な形態から数多くの分枝を持つ複雑な形態のものまで、非常に多種多様な神経細胞を含んでいます。 いずれのタイプの神経細胞もすべて神経幹細胞によって生み出されていると考えられています。しかし、これほど多様な神経細胞をひとつひとつ作り分けるメカニズムの全貌は未解明のままです。
私たちはこれまでに、神経幹細胞で一過的に発現するTemporal Factorsと呼ばれる転写因子群が神経多様性の創出に重要であることを報告してきました(Suzuki et al., 2013; 2016)。 発現スクリーニングにより、これから神経を生み出す若い幹細胞、何度か分裂した中間期の幹細胞、すでに多くの神経を生み出した古い幹細胞において、それぞれ特異的に発現がみられる遺伝子を同定しました。 それらの機能を解析した結果、それぞれの遺伝子が、生み出される神経のタイプの決定に関わっていることがわかりました。 つまり、神経幹細胞において発現する遺伝子が次々に変化することによって、生み出される神経の種類も変化し、結果として神経の多様性が作り出させると考えられます。
現在では、それぞれのTemporal Factorの発現の経時的な変化を制御するメカニズムやTemporal Factorが神経細胞のタイプを決定する分子機構などを研究しています。
正確な神経回路を構築する仕組み
神経細胞は生み出された後、自身のパートナーとなる細胞と結合して神経回路を形成しそれぞれの機能を果たします。 神経回路の構築には、神経細胞が生まれた場所から移動して自分のパートナーを発見・認識し結合する、という複数のステップが含まれています。 回路形成の制御は正しく機能する脳を形成するために非常に重要ですが、いずれの制御メカニズムにも不明な点が多く残されています。
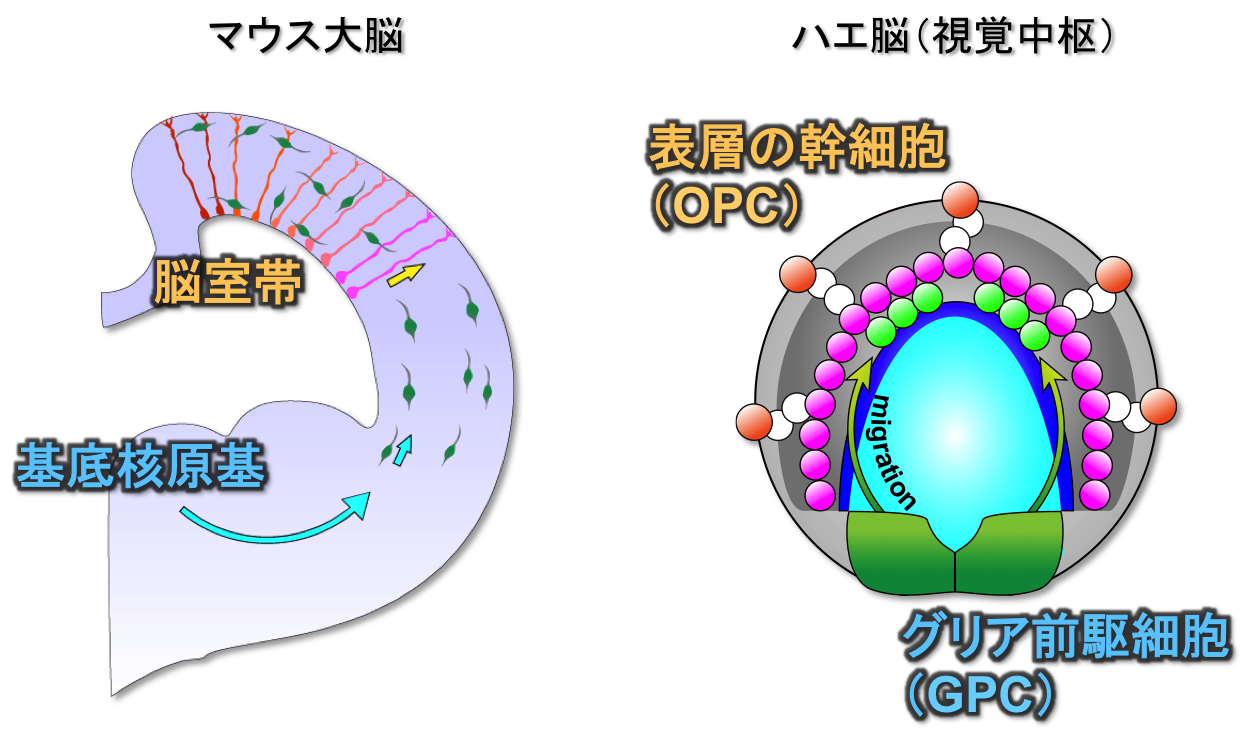
哺乳類の大脳皮質では、2つの神経供給源が知られており(脳室帯、基底核原基)、それぞれ興奮性神経、抑制性神経を産生しています。 これら由来の異なる神経細胞が相互作用することによって複雑な神経回路を作り上げています。 私たちは、このような由来の異なる神経細胞による回路形成がショウジョウバエでも見られる(OPC, GPC)ことを報告してきました(Suzuki et al., 2016)。 GPCで生まれた神経細胞は接線方向に移動することによってメダラに供給され、OPC由来の神経細胞と結合すると考えられます。
現在では、同じ情報処理回路に関与すると考えられている一組の神経(GPC由来神経とOPC由来神経の一種) に注目し、細胞移動からパートナー認識までのメカニズムを研究しています。